
Genetičko podrijetlo Hrvata
Dosadašnja istraživanja genetičkog podrijetla i rezultati genetičkih analiza omogućuju tumačenje genetičkog podrijetla Hrvata. Postoji samo nekoliko radova koji među analizama haplotipova na Y kromosomu sadrže i analize uzoraka iz Hrvatske. Od početnih radova: Semino i suradnici (koautor Dragan Primorac) 2000., Passarino i sur. 2001. i Chikhi i sur. 2002., genetika je kao znanost dosta napredovala. Kod ovakvih istraživanja nezaobilazno je proučavanje i znanstvenih radova koji se bave drugim narodima i područjima, jer omogućavaju razumijevanje geografskih ishodišta pojedinih haplotipova, njihovog razmještaja na Zemlji i širenja.
U istraživanju genetike razlikujemo 3 odvojena testiranja;
- DNK koja se nalazi na Y kromosomu i predstavlja izravnu mušku liniju, Y-DNK i koja obično služi za praćenje pravaca migracija (muškarci su se u pravilu više selili nego žene), a koja u hrvatskom primjeru prikazuje doseljavanje s područja Karpata i Ukrajine i u manjem dijelu autohtonost.
- DNK koja se nalazi u mitohondrijima i predstavlja izravnu žensku liniju, mtDNK koja je u hrvatskom primjeru u nekom europskom prosjeku.
- autosomalnu DNK koja predstavlja obilježja cijele populacije, uzima u obzir sve linije, maksimalno 2 na n-tu, pri čemu je n broj generacija (broj linija je realno manji, jer su se nakon n generacija i neki rođaci ženili međusobno)
Teritorij Hrvatske je naseljen od paleolitika, u neolitiku se tu nalaze kulture poput vučedolske, lasinjske, Sopota, Korena, impreso-keramičke, Danila i Hvara. U brončanom dobu dolazi do simbioze Protoindoeuropljana i neolitske kulture, što vodi nastajanju grupa naroda poput Ilira. S većim osvajanjima i seobama Kelta, Rimljana, Avara i na kraju Slavena/Hrvata u 6. stoljeću većinom se završavaju velike migracije. Dodatne selidbe stanovništva prouzročene su hrvatsko-otomanskim ratovima, hrvatskim ekonomskim migracijama u 19. i 20. st-u, svjetskim ratovima, kao i Domovinskim ratom 90-ih.
Ovo područje se brzo razvija i stalno se pojavljuju nova istraživanja, tako da je neke konačne zaključke prerano iznositi. U tekstu se izraz podrijetlo Hrvata koristi kao sinonim za podrijetlo populacije stanovništva s područja Hrvatske, jer je prema prema popisu stanovništva iz 2001. godine u populaciji stanovništva Republike Hrvatske bilo 89,65% Hrvata, uz znatan postotak nepoznate narodnosti. Međutim, Hrvati žive i izvan Hrvatske.
Treba razlikovati genetičko podrijetlo naroda od stvaranja političkih naroda/nacija (etnogeneze). Pojedini haplotipovi nisu povezani s etničkom pripadnošću. Autori radova s područja genetike daju im ponekad, radi lakšeg razlikovanja, imena prema narodima ili skupinama u kojima je otkriven njihov najveći postotak. Sva istraživanja ne koriste istu terminologiju i razlučivanje, pa su zaključci iz jednog istraživanja često upitni kad se pojavi novo istraživanje.
Početna istraživanja haplotipova pronađeni u uzorcima iz Hrvatske (po istraživanju Semino i sur. )
[uredi | uredi kôd]

Početna istraživanja, kao npr. Semino i sur., na 58 muškaraca iz Hrvatske nađene su sljedeće skupine kromosoma Y:
- haplotip I2 (Eu7): 44,83%
- haplotip R1a (Eu19): 29,31%
- "neolitski haplotipovi" E, J, G (Eu4, Eu9, Eu10, Eu11): 13,79%
- haplotip R1b (Eu18): 10,34%
- haplotip O (Eu16): 1,73
Ovakva genetička struktura slična je strukturama susjednih naroda jugoistočne Europe, iako postoje i određene razlike. Usporedbe s drugim narodima moguće je načiniti ako su istraživanja njihova genetičkog podrijetla načinjena po istoj metodi: analizom haplotipova. Ako se radi o starijim istraživanjima temeljenim na haplogrupama, može se samo izračunati sličnost podrijetla predaka uspoređivanih naroda.
Najuočljivija karakteristika prikazane genetičke strukture je visok postotak haplotipa Eu7. Kasnije se pokazalo da je hapolotip I2(a) porijeklom iz Ukrajine, odnosno da nije preživio ledeno doba u Dinaridima, nego da se prvo proširio preko Karpata, sudjelovao u slavenskoj etnogenezi tamo, a zatim skupa s R1a (a vjerojatno i s drugim haplogrupama u tragovima) naknadno doselio/vratio na ova područja.
Genetički, po izravnoj muškoj liniji, većina Hrvata (>85%) pripada jednoj od tri glavne europske haplogrupe;
- haplogrupa I 38%[1][2][3]-44%[4]
- haplogrupa R1a 27%[4]-34%[1][2][3]
- haplogrupa R1b 12.4%[4]-15%[1][2][3]
Dok manjina (>15%) većinom pripada haplogrupama;
Haplogrupa I se u Hrvata nalazi u 2 grupe;
- I2 (35%), karakteristična za istočni Jadran, Podunavlje i Ukrajinu od kuda potječe
- I1 (9%), karakteristična za Skandinaviju[4]

Od I2 grane, koja je tipična za slavenske populacije jugoistočne europe, a nalazi maksimum među Hrvatima Bosne i Hercegovine i južne Hrvatske (Like i Dalmacije), gdje prelazi i 50% ("efekt osnivača" na djelu)[3] kao npr. u Hercegovini (73%),[5] Dubrovniku (53%) and Zadru (60%),[6] kao i na južnim otocima poput Visa (44,6%),[6] Brača, Korčule (~55%), i Hvara (65%).[7] Rasprostranjenost te grupe je niža u gradovima na sjeveru poput Osijeka (27,6%) i zapadnim planinama poput Žumberka (18,2%)[6] i sjevernim otocima Cresu (3%) i Krku (27%).[5][1] I1 grana nije pronađena u Osijeku i među BiH Hrvatima, ali je imala maksimum od 8,9% u Dubrovniku.[6]

Od I2a haplogrupe najčešća je podgrana I2a1 (I-P37.2), odnosno I-M423 (nastala u razdoblju prije 18.400-14.000 godina), podskup I-Y3104 (nastala prije 14.000-11.400 godina), podskup I-L621 (nastao prije 11.400-6.500 godina), podskup I-CTS10936 (nastao prije 6.500-5.600 godina), podskup I-S19848 (nastao prije 5.600 godina), podskup I-CTS4002 (nastao prije 5.600-5.100 godina), podskup I-CTS10228 (nastao prije 5.100-3.400 godina), podskup I-Y3120 (nastao prije 3.400-2.100 godina), podskup I-S17250 (nastao prije 2.100-1.850 godina) > I-PH908 (nastao prije 1.850-1.700 godina),[8]
Najranije arheološko nalazište ove grupe do sada je Sungir 6 (730-850) u blizini grada Vladimira u Rusiji koje je pripadalo podgrupi I-S17250, podskup I-Y5596, podskup I-Z16971, podskup I-Y5595, podskup I-A16681 kluster,[9][10] kao i I-CTS10228 i I-Y3120 podgrana koje su nađene u dva vikinška uzorka iz Švedske (VK53) i Ukrajine (VK542) pretežno slavenskog porijekla od čega potonja pripada Glebu Svyatoslaviću, primcu Novgorodskom iz 11. stoljeća.[11]
Populacija s ovom haplogrupom je u Europu stigla s Bliskog istoka, otprilike prije 25-13 tisuća godina. To su ljudi iz Paleolitika i Mezolitika (lovci-sakupljači).
Podgrupa I-P37.2 ima veliku rasprostranjenost u RH i okolnim zemljama.[12][6] Međutim, u usporedbi sa starijim radovima koji su predlagali "autohtono porijeklo" haplogrupe I-P37 u Hrvatskoj,[nb 1] već je Battaglia et al. (2009) uočila najveću raznolikost i brojnost haplogrupe na prostoru Ukrajine, a Zupan et al. (2013) zaključio kako to sugerira dolazak u trenutku slavenske migracije iz pradomovine s prostora Ukrajine.[18]
Najnovije istraživanje koje je napravila O. M. Utevska (2017),[19] zaključilo je kako haplogrupski STR haplotip ima najveću raznolikost u Ukrajini, gdje se nalazi ancestralni STR marker "DYS448=20" koji čini "dnjeparsko-karpatski" kluster te derivirani "jugoistočno europski" kluster markera "DYS448=19" koji čini većinu I2 kod južnih Slavena. Taj "dinaridsko-podunavski kluster" također ima najveću raznolikost na području Ukrajine, ali ne i kod južnih Slavena gdje je najveći postotak što pokazuje na "efekt osnivača" na prostoru Dinarida, Podunavlja i Primorja.
Utevska je izračunala kako razdvajanje klustera i ekspanzija prema Jadranu se dogodila otprilike prije 2.860 ± 730 godina, što se podudara s razdobljem formacije i migracije Slavena (samim tim i doseljenja Hrvata),[19] odnosno, kluster predstavlja samo jedan SNP, I-PH908, imena I2a1a2b1a1a1c na ISOGG filogenetskom stablu (2018), a prema YFull YTree kalkulaciji imao je formaciju i TMRCA oko 3. stoljeća.[17]
Iako se smatra kako je I-621 mogla biti prisutna u Cucuteni-tripilijskoj kulturi,[20] do sada je samo pronađena haplogrupa G2a,[21] a druga grana I2a1a1-CTS595 je bila prisutna u badenskoj kulturi u Panonskoj nizini.[20][22] Iako su I-L621 i njezine podgrane dominantne kod novodoseljenih Slavena na području bivšeg Rimskog carstva, do sada nisu pronađene u arheogenetskim uzorcima iz rimskoga razdbolja i gotovo ih nema u suvremenoj populaciji Italije.[7]
Pronađena je u skeletnim ostacima doseljenika s artefaktima, ukazujući na vođe, kao i kod mađarskih osvajača panonske nizine iz kasnog 9. stoljeća, kao dio (slavenske) zapadne euroazijske-slavenske komponente suvremenih Mađara.[20][7] Prema radu Fóthi et al. (2020), rasprostranjenost ancestralnih podgrana poput I-CTS10228 u suvremenim populacijama ukazuje na naglo širenje s područja jugoistočne Poljske, uglavnom je vezano za Slavene (samim tim i doseljenja Hrvata), i "najveća demografska eksplozija se dogodila u Dinaridima i Podunavlju".[7]
Haplogrupe R1a i R1b su u Hrvata druga (34%) i treća (15,6%) haplogrupa po brojnosti u Hrvata, po istraživanju iz 2003.[1] Po istraživanju iz 2008. njihove vrijednosti su samo malo manje.[4] Haplogrupa R1a (R-M17), odnosno njena R1a1a1 podgana se u Hrvatskoj većinom nalazi u 2-3 podgrane;
- R1a#R-M458 (R1a1a1b1a1 ili R-M558) koja je većinska
- R1a#R-Z282 (R1a1a1b1a, u podgranama R-M458 i R-Z282) koja je rjeđa[6]
Najveća frekvencija R1a je nađena među Hrvatima Osijeka (39%){sfn|Battaglia et al.|2008}}, Žumberka[6] i na sjevernim otocima Krku (37%) i Cresu (56,5%)[1][6] gdje su slične i rezultatima drugih Slavena, poput Slovenaca, Čeha i Slovaka. Frekvencija je niža u Zadru (4%) i Dubrovniku (13,4%), kao i na južnim otocima Hvaru (8%), Visu (17%), Korčuli (20%) i Braču (25%)[1][6] U BiH Hrvata, frekvencija je slična onoj drugih južnih Slavena (12%).[4]
Haplogrupa R1b, koja se u Hrvatskoj nalazi u više podgrana, ima najveći postotak u Hrvata s Dugog otoka (25%) i Krka (16,2%)[1][6], u Žumberku je bila 11,3%[6], dok je na južnim otocima, gradu Dubrovniku (3,9%) i BiH Hrvatima jako rijetka (1-6%)[1][4][6], dok u Osijeku nije uopće nađena.[4]
Ove dvije haplogrupe (R1a i R1b) su povezane s indoeuropskim seobama iz Euroazije prije kojih 5 tisuća godina[12], pri čemu se R1a (uz I2a) posebno veže sa seobom Slavena.[3][6] Njihove frekvencije obrnuto zrcale razmještaj haplogrupe I2a[12] i najjače su u sjevernim, zapadnim i istočnim djelovima Hrvatske[1][4], čime pokazuju "efekt osnivača", odnosno doseljenje Hrvata kroz primarno te dvije haplogrupe (R1a i I2a).
R-M558 podgrana CTS1211 je također pronađena među mađarskim osvajačima što ukazuje na miješanje i asimilaciju Slavena i Hrvata među Mađarima.[20]

Od haplogrupe E u Hrvata je najčešća podgrana E-V66#E-V13|E1b1b1a1b-V13 (6,7%), a i E1b1b1a3-M149 i E-M123|E1b1b1c-M123 su isto nađene u malim postocima (1,1%).[5] E-V13 je tipična za populaciju jugoistočne Europe, ima vrhunac među Kosovskim Albancima (44%), ali je isto visoka i među Makedoncima, Bugarima, Grcima, Rumunjima i Srbima.[3] Najveća frekvencija je nađena u Žumberku (18,2%) [6] i Osijeku (10,3%)[4], na otocima Dugi otok (15,9%) i Ugljanu (13,2%), kao i južnim otocima Visu (23,4%) i Mljetu (15,4%).[6] Na sjevernim otocima Cresu (3%)[6] i Krku (6,8%) je bila slična drugim južnim otocima (3,7-4,3%).[1] U BiH Hrvata je njena frekvencija ista kao i u RH Hrvata (8,9%).[5]
Podgrane J1 su rijetke u Hrvatskoj, dok je J2 veća kod Hrvata u Hrvatskoj, najveći postotak imaju Osijek (10,2%) i središnji otoci Pašman (16,6%) i Ugljan (10,2%) kao i sjeverni otoci Krk (10,8%) i Cres (14,1%)[1][6][5] nego BiH Hrvati (1,1%).[5]
Podgrana G2a-P15 je nađena kod RH i BiH Hrvata u niskom postotku (1,1%),[5] ali je vrhunac našla u sjeveroistočnom gradu Osijeku (13,8%)[5] i južnim otocima Mljetu (15,4%), Korčuli (10,4%), Braču(6%), kao i sjevernom otoku Cresu (7%).[1][6]
Haplogrupe E i J su povezane s neolitskim migracijama prvih poljoprivrednika iz Anatolije/Bliskog istoka koji su sa sobom doveli udomaćene životinje i poljoprivredu. Mutacija haplogrupe E (V13) se vjerojatno dogodila na Balkanu, ali ne ranije od 8-10 tisuća godina. Ove haplogrupe pokazuju gradaciju od sjevera prema jugu, gdje ih je više.[12] Haplogrupa G je mogla biti prisutna u Europi i prije ovih seoba, za vrijeme ledenog doba, ili su neke njezine podgrane došle s ranim poljoprivrednicima.[23]
Haplogrupa N je rijetka u Hrvatskoj (2,2%).[5] Česta je na Dalekom istoku, poput Sibira i Kine, a u Europi među Fincima (60%) i u Baltičkim zemljama (45%). Neobično za europske populacije, još jedna središnjoazijsko-sibirska grupa P, poput haplogrupe Q je nađena u visokoj frekvenciji na otocima Hvaru (14%) i Korčuli (6%).[1]
U radu iz 2014. iz troje neolitskih uzoraka Starčevačke kulture iz Vinkovaca, dvoje je pripadalo G2a-P15 Y-DNK grupi, a jedan I2a1-P37.2, što bi pokazalo G2a kao potencijalne predstavnike širenja poljoprivrede s Bliskog istoka u Europu, dok bi I2a bio Mezolitski ostatak u Europi.[24] U radu iz 2018., iz 10 od 17 uzoraka iz Hrvatske je uspješno izvučena Y-DNA; dvoje iz središnjeg Neolitika (6005-5786 PK) iz špilje Zemunica koji su pripadali C1a2 i E1b1b1a1b1, ranog Neolitika Starčevačke kulture (5837-5659 PK) iz Belog Manastira-Popove zemlje haplogrupi C, i ranog Neolitika Impresso-keramičke kulture (5641-5560 BCE) iz Kargadura koji su pripadali G2a2a1, 2 srednjeg Neolitska u Sopotu (52007-4546 PK) uzorka iz Osijeka koji su pripadali G2a2a1 i J2a1, kasno neolitski Sopot (4790-4558 BCE) iz Belog Manastira-Popove zemlje haplogrupi I, 2 iz Vučedola (2884-2582 BCE) iz Belog Manastira-Popove zemlje i Vučedola koji su pripadali R1b1a1a2a2 i G2a2a1a2a, te ranom Brončanom dobu (1631-1521 BCE) iz Velikog Vanika, koji pripada J2b2a.[25]
U radu iz 2012. je provedena dotad najopširnije istraživanje genetike u Hrvata. Analizirano je 17 markera na Y kromosomu, a predviđanja su ostvarena s 90% sigurnošću.[26] Populacija je podijeljena na 5 cjelina, pri čemu je vidljiva velika homogenost ukupnog materijala, kao i po cjelinama (s blagom iznimkom dijela južne Hrvatske). Dodatno na veliku ukupnu homogenost primjetan je i lagani gradijent od sjevera prema jugu (i istoku i zapadu) od srednjeeuropskog (germanskog, slavenskog, mađarskog) klustera prema jugoistočnoeuropskog (južnotalijanskog, grčkog, balkanskog).[26]
| Population | Samples | Source | I2a(xI2a1) | R1a | E1b1b1-M35 | R1b | I1 | J2b | G2a | H | J2a1h | J1 | J2a1b | E1b1a1-M2 | G2c | I2a1 | I2b1 | I2b(xI2b1) | J2a1-bh | L | N | Q | T |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Overall Croatia | 1,100 | Mršić et al. (2012) | 37.7 | 22.1 | 10.6 | 7.9 | 5.8 | 3.7 | 2.7 | 1.8 | 1.2 | 1.1 | 1 | <1 | <1 | <1 | <1 | <1 | <1 | <1 | <1 | <1 | <1 |
| Central Croatia | 220 | Mršić et al. (2012) | 31.8 | 23.6 | 11.8 | 10.4 | 5 | 5 | 3.6 | 1.3 | 0.4 | 2.2 | 0.9 | 0 | 0 | 0.4 | 0.9 | 0.9 | 0 | 0.4 | 0.9 | 0 | 0 |
| North Croatia | 220 | Mršić et al. (2012) | 25.4 | 29.1 | 10.9 | 10.4 | 4.1 | 5 | 3.1 | 5 | 0.4 | 0 | 0.4 | 0.4 | 0 | 0 | 2.2 | 0.9 | 0 | 0.4 | 0.4 | 0 | 1.3 |
| East Croatia | 220 | Mršić et al. (2012) | 40 | 18.6 | 11.3 | 8.2 | 5.9 | 2.7 | 1.8 | 0.9 | 2.2 | 2.7 | 1.3 | 0 | 0 | 0 | 0.4 | 0 | 0.4 | 0 | 0.9 | 1.8 | 0.4 |
| West Croatia | 220 | Mršić et al. (2012) | 36.8 | 20 | 12.7 | 5.9 | 8.6 | 3.2 | 3.2 | 1.8 | 1.8 | 0.4 | 1.8 | 0 | 0.4 | 0 | 0.4 | 0 | 0.4 | 0 | 0.9 | 0.4 | 0.9 |
| South Croatia | 220 | Mršić et al. (2012) | 54.5 | 19.1 | 6.3 | 4.5 | 5.4 | 2.7 | 1.8 | 0.4 | 0.9 | 0 | 0.4 | 0 | 0 | 0 | 0.4 | 0 | 0.9 | 0.4 | 0 | 1.3 | 0.4 |
Haplotipovi pronađeni u uzorcima iz Hrvatske, BiH, Srbije i Crne Gore (prema podatcima s Eupedije)
[uredi | uredi kôd]Haplotipovi pronađeni u uzorcima iz Hrvatske, BiH, Srbije i Crne Gore (prema podatcima s Eupedije)[27]
- I1: predgermanski (sjevernjački)
- I2b: predkelto-germanski
- I2a1: sardinski, pirenejski
- I2a2: dinarski, dunavski
- N1c1: uralofinski, baltički, sibirski
- G2a: kavkaski, grčko-anatolski
- E1b1b: sjevernoafrički, bliskoistočni, balkanski
- T: srednjoistočni, istočnoafrički
- R1a: balto-slavenski, germanski, indo-iranski
- R1b: italo-keltski, germanski, hetitski, armenski, toharski
- J1: kavkaski, mezopotamski, semitski (arapski, židovski)
- J2: grčko-anatolski, mezopotamski, kavkaski
| Haplogrupa | I1 | I2*/I2a | I2b | R1a | R1b | G | J2 | J*/J1 | E1b1b | T | Q | N |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hrvatska | 5,5 | 37 | 1 | 24 | 8,5 | 2,5 | 6 | 1 | 10 | 0,5 | 1 | 0,5 |
| BiH | 4,5 | 50,5 | 0 | 18 | 3,5 | 1,5 | 5,5 | 1 | 11,5 | 0,5 | 0,5 | 1 |
| Bošnjaci iz BiH | 4 | 53,5 | 0 | 18 | 3,5 | 1,5 | 5 | 0,5 | 9 | 0 | 0,5 | 1 |
| Hrvati iz BiH | 0 | 71 | 2 | 12 | 2 | 1 | 1 | 0 | 9 | 0 | 0 | 0 |
| Srbi iz BiH | 7,5 | 32,5 | 0,5 | 20 | 3,5 | 1 | 8,5 | 3,5 | 17,5 | 1 | 0 | 1 |
| Srbija | 8 | 34 | 0,5 | 18 | 6 | 2,5 | 9 | 1 | 15 | 0,5 | 1 | 2,5 |
| Crna Gora | 6 | 29,5 | 1,5 | 7,5 | 9,5 | 2,5 | 9 | 0,5 | 27 | 0 | 2 | 1,5 |
-
 Y-haplogrupe u Hrvatskoj
Y-haplogrupe u Hrvatskoj -
 Y-haplogrupe Hrvata iz BiH
Y-haplogrupe Hrvata iz BiH -
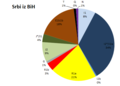 Y-haplogrupe Srba iz BiH
Y-haplogrupe Srba iz BiH -
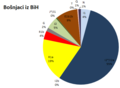 Y-haplogrupe Bošnjaka iz BiH
Y-haplogrupe Bošnjaka iz BiH




Genetički, preko izravne majčinske linije većina (>65%) hrvatske populacije iz RH pripada jednoj od tri od jedanaest glavnih europskih mtDNK grupa;
- haplogrupi H (45%)
- haplogrupi U (17,8-20,8%)
- haplogrupi J (3-11%)[28]
Haplogrupa H je najčešća grupa, koja maksimalnu frekvenciju pokazuje na otocima Korčuli (60,2%) i Mljetu (73,5%)[29], a minimalnu na otocima Cresu (27,7%)[30], i Hvaru (27.8%).[28] Riječ je o dominantnoj europskoj haplogrupi.[29] Povećana frekvencija podgrupe H1 na Mljetu (30,9%), inače rijetke u ostalim istraživanjima je standardni primjer "efekta osnivača" - selidbe iz najbližeg obalnog područja i mikroevolucijskom širenju po otoku.[29]
Haplogrupa U (18,85% in 2014)[31] je većinom zastupana svojom podgrupom U5, koja je druga najraširenija haplogrupa s frekvencijom 11,6% na kopnu i 10,4% na obali, sa sličnim frekvencijama na otocima Braču (8,6%), Krku (11,3%), i Hvaru (14,8%), dok je najniža na Korčuli (2%).[28]. Ukupno je haplogrupa U, sa svim svojim podgranama (poput U5), najčešća u gradu Dubrovniku (19,9%) i otocima Lastovu (23,5%) i Cresu (29,4%).[30][29] Ona je najstarija europska haplogrupa (nastala prije oko 50 tisuća godina), a njezina podgrupa U5 čini većinu unutar nje u europi. Veliki postotak podgrane U4 na Lastovu (11,8%) ukazuje na "efekt osnivača".[29]
Haplogrupa J je treća najčešća, s 11,9% na kopnu, ali samo 3,1% na obali, međutim neki otoci su imali više frekvencije od obalnih populacija; Korčula (6,1%), Brač (8,6%), Krk (9%), Hvar (9,3%)[28], a maksimum ima na planinama Žumberka (12,34%)[31] i Lastovu (19,6%),[29] dok je na Cresu gotovo potpuno odsutna.[30]
Od haplogrupe T, podgrupa T2 ima sličnu učestalost (3,1-5-8%) in na obali i na kopnu, kao i među otočnom populacijom s maksimumom na Hvaru (12%)[28][30], no i minimumu na Mljetu (1.5%), Lastovu (3.9%) i u Dubrovniku (2,3%).[29]
Haplogrupa K ima prosječnu frekvenciju 3,6% na kopnu i 6,3% na obali, odsutna je na otoku Lastovu[29] i ima najnižu frekvenciju na Cresu (3,36%)[30] i Hvaru (3,7%), a najvišu na Braču (9,5%).[28]
Haplogrupa V je mlađa podgrana haplogupe H[29] i ima otprilike iste minimume i maksimume (4-5,7%) i na obali i na kopnu, s iznimkom na Korčuli (8,2%)[28] kao i miminumima na Mljetu (1,5%), Lastovu (2%) i Dubrovniku (3,3%).[29]
Haplogrupa W ima frekvenciju između 2,2 i 4,2% na kopnu i obali, a na otocima varira između 1,9 i 3,1%, s iznimkom na Krku (7,5%)[28] i Cresu (12,6%).[30] Na otocima Mljetu i Lastovu je između 4,4 i 5,9%, dok je u Dubrovniku skoro potpuno odsutna (0,6%).[29]
Druge mtDNKa s primjetljivim vrhovima su:
- HV podgrupe s nižim frekvencijama na kopnu i obali (0,4-2,1%), ali u prosjeku (4,1-4,6%) na otocima, i maksimumu u Dubrovniku (7.7%)[29] i na Braču (10.5%).[28]
- Haplogrupa N1a na Cresu (9.24%) je najsjeverniji nalaz te grupe dosad u Europi, i pokazuje na relativno nedavan "efekt osnivača"[30]. To je karakteristična haplogrupa ranih poljoprivrednika.[24]
- Haplogrupa F koja je skoro pa potpuno odsutna (samo 0,2-0,4%) na kopnu, ali ima maksimum od 8,3% na Hvaru.[28][31]
- Haplogrupa I na Krku (11,3%), s podgrupama koje su se odvojile u Ledenom dobu.[32]
Rad iz 2011. na 27 ranosrednjovjekovnih ostataka kostura iz Naklica blizu Omiša u južnoj Dalmaciji je pokazalo da ih je 67% pripadalo haplogrupi H, 18% J, 11% U5 i 4% HV.[33] Rad iz 2015. na srednjovjekovnim ostacima kostura u Šopotu (14-15. st) i Ostrovici iz 9. stoljeća u sjevernoj Dalmaciji su potvrdili kontinuitet u tom razdoblju, pokazujući iste haplogrupe (H, J, U) i u srednjovjekovnoj i današnjoj populaciji.[34] Rad iz 2014. na muškom kosturu iz Splita iz kasnorimskog razdoblja (otprilike 1760±80 godina) je pokazalo da je isti pripada haplogrupi H.[35]
U radu iz 2014. koji je obradio i y i mt DNKa, jedan mezolitski uzorak datiran 6080-6020 p.K. iz Vele Špilje blizu Vele Luke na otoku Korčuli je pripadao mtDNK haplogrupo U5b2a5 uobičajenoj za populaciji lovaca-sakupljača, dok je drugih jedanasest neolitskih uzoraka iz Starčevačke kulture uzoraka, datiranih 6000-5400 p.K. iz Vinkovaca pripadalo haplogrupama J1c, K1a, T2b, HV0, K, V, V6, što otkriva sličnu mtDNK raznolikost i liniju potomstva između ranih populacija Panonskog bazena s populacijom Srednje Europe koja je pripadala Kulturi Linearnotrakaste keramike, uz prorijeđivanje mezolitskog mtDNKa ostatka.[24]
Preliminarni rezultati rada iz 2016. o mtDNK, koji bi trebali uključiti 30 uzoraka iz Neolitika i 5 uzoraka iz kasnog brončanog doba, na 5 antičkih kostura(3 neolitska iz Kulture Linearnotrakaste keramike iz špilje Zemunica blizu Biska, 1 srednjoneolitska Danilske kulture iz Zidane špilje blizu Staničića Žumberačkih, 1 sredno/kasno brončano dobna iz Špilje Jazinka blizu Nečvena) pokazuju haplogrupe K2 i K1b1a, H1e/H41, H1b za neolitske uzorke slične onima ranih europskih poljoprivrednika, modernih Sardinijanaca i južni Europljana, dok haplogrupe HV ili H4 iz Brončanog doba pokazuju sličnost modernom stanovništvu Hrvatske i jugoistočne Europe, ali bez jasnih dokaza koji pokazuju poveznicu s migracijom Indoeuropljana.[36]
Rad iz 2018. o 17 uzoraka iz Hrvatske;
- Mezolitske (7308-7027 p.k.) iz Vela Špilja (U5b2b)
- tri s početka Neolita (6005-5751 p.k.) iz Špilje Zemunica (H1, K1b1a i N1a1)
- ranoneolitskog Starčeva (5873-5659 p.k.) iz Beli Manastir-Popova zemlja (U8b1b1)
- dva ranoneolitska iz Linearnotrakaste keramike (5670-5560 p.k.) iz Kargadura (H5a, H7c)
- dva srednjoneolitska iz Sopota (5207-4546 p.k.)iz Osijeka (U5a1a2 i H10)
- dva kasnoneolitska iz Sopota (4790-4536 p.k.) iz Osijeka (U5b2b i N1a1)
- eneolitskog iz Radovanaca (3710-3360 p.k.) (J1c2)
- tri iz Vučedol (3000-2582 p.k.) iz Beli Manastir-Popova zemlja i Vučedol (T2e, T2c2 i U4a)
- rano-srednjo brončani (1631-1521 p.k.) iz Velikog Vanika (I1a1)
- kasno brončani (805-761 p.k.) uzorak iz špilje Jazinka koji su pripadali HV0e.[25]
Desetljećima su hrvatski otoci bili proučavani zbog njihove izolacije preko koje se lakše uočavaju mikroevolucijski procesi radi boljeg razumijevanja evolucijskih silnica poput genetičkog drifta(specifičnog genetičkog izražaja), "efekta osnivača" i populacijskih "uskih grla" (smanjivanja veličine populacije) koji su oblikovali modernu populaciju. Dosadašnji rezultati pokazuju da je genetski tok i doseljavanje žena na otoke bilo ograničeno.[29]
Slični procesi (umjerene genetičke izoliranosti) se mogu primijeniti i na kopnena područja brdskih regija poput Žumberka, gdje su oni imali labavu vezu s regijom nastanka Uskoka (južna RH i BiH) ili s njihovim najbližim sadašnjim susjedima (sjever RH i Slovenija).[31]
Na primjeru populacije otoka Krka, analiza visoke rezolucije mtDNK pokazuje dokaze da su naselja Omišalj, Vrbnik i Dobrinj povezana u grupu ranog slavenskog naseljavanja, dok su područja Poljica i Dubašice bliža drugoj grupi slavenskog i vlaškog/morlačkog doseljavanja iz velebitskog zaleđa u 15. stoljeću.[32] Na primjeru populacije Mljeta može se povezati napraviti demografsko-povijesni lanac događaja kao upotreba otoka kao mjesta za karantenu, uz često sparivanje unutar zajednice poput Visa i Lastova radi manjka genetičke raznolikosti.[29]
U analizi iz 2004, uspoređivali su se grupe populacije otoka Hvara, Krka i Brača s drugom grupom hrvatskog kopna i obale, dok je otok Korčula bio izdvojen radi velike frekvencije haplogrupe H.[28]
U analizi iz 2009. otočne populacije Krka, Ugljana, Brača i Hvara su se zbili skupa, pokazujući da imaju bliske majčinske linije, s Visom blizu grupe, dok su Cres (većina U2, W, N1a) i Rab] (U4, H6, J1c) imali izdvojene pozicije iz grupe i međusobno.[30]
U analizi iz 2014. populacije istočne i južne Hrvatske su se zbile skupa s BiH, dok je zapadna i sjeverna Hrvatska naginjala više prema Sloveniji. Kako slovenska populacije nije dio Jugoistočno-europske grupe, razmatrao se mogući unos drugačijih seoba Slavena u srednjem vijeku.[31]
Prema analizi iz 2014. godine, hrvatska populacija pokazuje načelnu srodnost drugim populacijama južnih Slavena. Genetska slika Hrvata i Bošnjaka je bliža genetskoj slici srednje Europe i preklapa se s genetskom slikom Mađara.[37] Prema analizi iz 2015. godine, grupirana je zapadna grupa Južnih Slavena u usporedbi s istočnom grupom u kojoj se nalaze Bugari i Makedonci, sa Srbima između. Zapadna grupa pokazuje sličnosti prema Mađarima, Česima i Slovacima, dok je istočna grupa srodna Rumunjima i u nekoj mjeri Grcima. Hrvatska populacija većinom dijeli zajedničke pretke s istočnim, zapadnim i južnim Europljanima i nema skoro nikakav odnos prema izoliranim sardinijskim i baskijskim populacijama.[37][38] Prema analizi iz 2016. svi od 176 uzoraka s otoka Visa su potvrdili izoliranost otočne populacije.[39]
-
 Admixture analysis of autosomal SNPs in a global context on the resolution level of 7 assumed ancestral populations per Kovačević et al. (2014)
Admixture analysis of autosomal SNPs in a global context on the resolution level of 7 assumed ancestral populations per Kovačević et al. (2014) -
 Admixture analysis on the resolution level of 6 assumed ancestral populations per Kushniarevich et al. (2015)
Admixture analysis on the resolution level of 6 assumed ancestral populations per Kushniarevich et al. (2015) -
 Genetic structure within European context according to three genetic systems atDNA (A), YDNA (B) and mtDNA (C) per Kushniarevich et al. (2015)
Genetic structure within European context according to three genetic systems atDNA (A), YDNA (B) and mtDNA (C) per Kushniarevich et al. (2015)



Izvore u ovom članku treba wikipedizirati. (Rasprava) |
- ↑ SNP I-P37 se formirao prije 20 tisuća godina i imao je najnovijeg zajedničkog pretka prije 18 tisuća godina prema YFull,[13] being too old and widespread as an SNP for argumentation of autochthony as well the old research used outdated nomenclature. According to "I-P37 (I2a)" project at Family Tree DNA, the divergence at STR marker DYS448 20 > 19 is reported since 2007,[14] while the SNP which defines the STR Dinaric-South cluster, I-PH908, is reported since 2016.[15] The SNP I-PH908 at ISOGG phylogenetic tree is named as I2a1a2b1a1a1c,[16] while formed and had TMRCA approximately 1,800 YBP according to YFull.[17]
- ↑ a b c d e f g h i j k l m n Barać et al. 2003. Pogreška u predlošku sfn: ne postoji izvor s oznakom: CITEREFBarać_et_al.2003 (pomoć)
- ↑ a b c Rootsi et al. 2004. Pogreška u predlošku sfn: ne postoji izvor s oznakom: CITEREFRootsi_et_al.2004 (pomoć)
- ↑ a b c d e f Peričić et al. 2005. Pogreška u predlošku sfn: ne postoji izvor s oznakom: CITEREFPeričić_et_al.2005 (pomoć)
- ↑ a b c d e f g h i j k l m n Battaglia et al. 2008. Pogreška u predlošku sfn: ne postoji izvor s oznakom: CITEREFBattaglia_et_al.2008 (pomoć)
- ↑ a b c d e f g h i Battaglia, Vincenza; Fornarino, Simona; Al-Zahery, Nadia; Olivieri, Anna; Pala, Maria; Myres, Natalie M; King, Roy J; Rootsi, Siiri; Marjanovic, Damir; Primorac, Dragan; Hadziselimovic, Rifat. Lipanj 2009. Y-chromosomal evidence of the cultural diffusion of agriculture in southeast Europe. European Journal of Human Genetics (engleski). 17 (6): 820–830. doi:10.1038/ejhg.2008.249. ISSN 1018-4813. PMC 2947100. PMID 19107149CS1 održavanje: format PMC-a (link)
- ↑ a b c d e f g h i j k l m n o p q r Šarac et al. 2016. Pogreška u predlošku sfn: ne postoji izvor s oznakom: CITEREFŠarac_et_al.2016 (pomoć)
- ↑ a b c d Fóthi, E.; Gonzalez, A.; Fehér, T. 2020. Genetic analysis of male Hungarian Conquerors: European and Asian paternal lineages of the conquering Hungarian tribes. Archaeological and Anthropological Sciences. doi:10.1007/s12520-019-00996-0
- ↑ I-PH908 YTree v8.06.01. YFull.com. 27. lipnja 2020. Pristupljeno 17. srpnja 2020.
- ↑ Sikora, Martin; Seguin-Orlando, Andaine; Sousa, Vitor C; Albrechtsen, Anders; Korneliussen, Thorfinn; Ko, Amy; Rasmussen, Simon; Dupanloup, Isabelle; Nigst, Philip R; Bosch, Marjolein D; Renaud, Gabriel; Allentoft, Morten E; Margaryan, Ashot; Vasilyev, Sergey V; Veselovskaya, Elizaveta V; Borutskaya, Svetlana B; Deviese, Thibaut; Comeskey, Dan; Higham, Tom; Manica, Andrea; Foley, Robert; Meltzer, David J; Nielsen, Rasmus; Excoffier, Laurent; Mirazon Lahr, Marta; Orlando, Ludovic; Willerslev, Eske. 2017. Ancient genomes show social and reproductive behavior of early Upper Paleolithic foragers. Science. 358 (6363): 659–662. doi:10.1126/science.aao1807. PMID 28982795
- ↑ I-A16681 YTree v8.06.01. YFull.com. 27. lipnja 2020. Pristupljeno 17. srpnja 2020.
- ↑ Margaryan, Ashot. 17. srpnja 2019. Population genomics of the Viking world. bioRxiv 10.1101/703405 journal zahtijeva
|journal=(pomoć) - ↑ a b c d Primorac et al. 2011. Pogreška u predlošku sfn: ne postoji izvor s oznakom: CITEREFPrimorac_et_al.2011 (pomoć)
- ↑ I-P37 YTree v6.07.08. YFull.com. 10. studenoga 2018. Pristupljeno 11. studenoga 2018.
- ↑ I2a Y-Haplogroup - Results: I2a2a-Dinaric. Family Tree DNA. Pristupljeno 11. studenoga 2018..
Ken Nordtvedt has split I2a2-M423-Dinaric into Din-N and Din-S. Din-N is older than Din-S. N=north of the Danube and S=south of the Danube River ... May 8, 2007: Dinaric I1b1 and DYS 448. DYS448 19 for S and 20 for N.
- ↑ Bernie Cullen. 23. rujna 2016. The Dinaric-South cluster is defined by the SNP PH908. i2aproject.blogspot.com. Blogger. Pristupljeno 11. studenoga 2018.
- ↑ Y-DNA Haplogroup I and its Subclades - 2018. ISOGG. 1. studenoga 2018. Pristupljeno 11. studenoga 2018.
- ↑ a b I-PH908 YTree v7.05.00. YFull.com. 9. lipnja 2019. Pristupljeno 21. lipnja 2019.
- ↑ Zupan et al. 2013. Pogreška u predlošku sfn: ne postoji izvor s oznakom: CITEREFZupan_et_al.2013 (pomoć)
- ↑ a b Utevska 2017, str. 219–226, 302. Pogreška u predlošku sfn: ne postoji izvor s oznakom: CITEREFUtevska2017 (pomoć)
- ↑ a b c d Neparáczki, Endre; Maróti, Zoltán. 2019. Y-chromosome haplogroups from Hun, Avar and conquering Hungarian period nomadic people of the Carpathian Basin. Scientific Reports. Nature Research. 9 (16569): 16569. doi:10.1038/s41598-019-53105-5. PMC 6851379. PMID 31719606
- ↑ Mathieson, Iain. 21. veljače 2018. The Genomic History of Southeastern Europe. Nature. 555 (7695): 197–203. doi:10.1038/nature25778. PMC 6091220. PMID 29466330
- ↑ Lipson, Mark. 2017. Parallel ancient genomic transects reveal complex population history of early European farmers. Nature. 551: 368–372. doi:10.1038/nature24476
- ↑ Primorac, Dragan; Marjanović, Damir; Rudan, Pavao; Villems, Richard; Underhill, Peter A. Lipanj 2011. Croatian genetic heritage: Y-chromosome story. Croatian Medical Journal. 52 (3): 225–234. doi:10.3325/cmj.2011.52.225. ISSN 0353-9504. PMC 3118711. PMID 21674820CS1 održavanje: format PMC-a (link)
- ↑ a b c Szécsényi-Nagy et al. 2015. Pogreška u predlošku sfn: ne postoji izvor s oznakom: CITEREFSzécsényi-Nagy_et_al.2015 (pomoć)
- ↑ a b Mathieson et al. 2018. Pogreška u predlošku sfn: ne postoji izvor s oznakom: CITEREFMathieson_et_al.2018 (pomoć)
- ↑ a b Mršić, Gordan; Gršković, Branka; Vrdoljak, Andro; Popović, Maja; Valpotić, Ivica; Anđelinović, Šimun; Stenzl, Vlastimil; Ehler, Edvard; Urban, Ludvik; Lacković, Gordana; Underhill, Peter. Srpanj 2012. Croatian national reference Y-STR haplotype database. Molecular Biology Reports (engleski). 39 (7): 7727–7741. doi:10.1007/s11033-012-1610-3. ISSN 0301-4851
- ↑ Maciamo. Eupedia. Eupedia (engleski). Pristupljeno 4. rujna 2022.
- ↑ a b c d e f g h i j k Cvjetan et al. 2004. Pogreška u predlošku sfn: ne postoji izvor s oznakom: CITEREFCvjetan_et_al.2004 (pomoć)
- ↑ a b c d e f g h i j k l m n Šarac et al. 2012. Pogreška u predlošku sfn: ne postoji izvor s oznakom: CITEREFŠarac_et_al.2012 (pomoć)
- ↑ a b c d e f g h Jeran et al. 2009. Pogreška u predlošku sfn: ne postoji izvor s oznakom: CITEREFJeran_et_al.2009 (pomoć)
- ↑ a b c d e Šarac et al. 2014. Pogreška u predlošku sfn: ne postoji izvor s oznakom: CITEREFŠarac_et_al.2014 (pomoć)
- ↑ a b Havaš Auguštin et al. 2012. Pogreška u predlošku sfn: ne postoji izvor s oznakom: CITEREFHavaš_Auguštin_et_al.2012 (pomoć)
- ↑ Ljubković et al. 2011. Pogreška u predlošku sfn: ne postoji izvor s oznakom: CITEREFLjubković_et_al.2011 (pomoć)
- ↑ Bašić et al. 2015. Pogreška u predlošku sfn: ne postoji izvor s oznakom: CITEREFBašić_et_al.2015 (pomoć)
- ↑ Sutlović, Davorka; Ljubković, Jelena; Stipišić, Angela; Knezović, Zlatka; Drmić-Hofman, Irena; Andjelinović, Šimun. 30. lipnja 2014. Skeletal Remains from Late Roman Period: »As Old as Diocletian's Palace«. Collegium antropologicum (engleski). 38 (2): 749–754. ISSN 0350-6134
- ↑ Fernandes et al. 2016. Pogreška u predlošku sfn: ne postoji izvor s oznakom: CITEREFFernandes_et_al.2016 (pomoć)
- ↑ a b Kovačević et al. 2014. Pogreška u predlošku sfn: ne postoji izvor s oznakom: CITEREFKovačević_et_al.2014 (pomoć)
- ↑ Delser et al. 2018. Pogreška u predlošku sfn: ne postoji izvor s oznakom: CITEREFDelser_et_al.2018 (pomoć)
- ↑ Jeroncic et al. 2016. Pogreška u predlošku sfn: ne postoji izvor s oznakom: CITEREFJeroncic_et_al.2016 (pomoć)
- Marijana Peričić, et al.: Review of Croatian Genetic Heritage as Revealed by Mitochondrial DNA and Y Chromosomal Lineages
 (PDF), Croat Med J 2005;46(4):502-513 (eng.)
(PDF), Croat Med J 2005;46(4):502-513 (eng.) - Dragan Primorac, Damir Marijanović, Pavao Rudan, Richard Villems, Peter Underhill Y kromosom i genetsko podrijetlo Hrvata
- Eupedia
| |||||||||||||||||||||||||||||||||||||||||||||||||||||
